Cross section of a hair
Rats have a wide variety of coat textures. From the smooth, standard coat to different types of wavy coat, to no coat at all. Where do these coat differences come from? How are hairs made, and how does variation in hair production lead to different textures of hair?
Disclaimer: I am a biologist, not a rat breeder, and my interest in rat coats stems from my fascination with the underlying biology. Therefore, this is not an article about rat standards! If you are seeking rat color standards, descriptions, and their associated Mendelian notations, check out one of the many websites of fancy rat organizations such as AFRMA, RMFE, NFRS, and AusRFS to name just a few.
The information on this page is based on information available in the scientific literature on rat coat types. This literature therefore deals with laboratory rats, not pet rats!
A word about coat names... rat coat names may vary between different countries and rat organizations. The same coat may be called by different names, and a particular name may refer to different coats, depending on who you talk to. Also, a particular coat may have a number of possible underlying causes, so use caution when extrapolating from this document to your pet rats. By and large, I have tried to steer clear of rat standards and "official" coat names, and have instead focused on my area of interest: the biology of coat production.
|
|
|
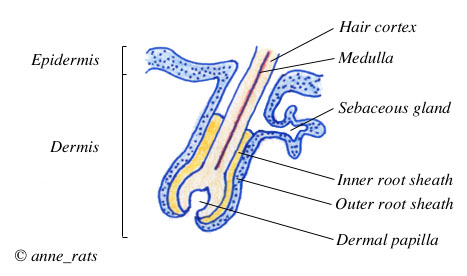
Cross section of a hair |
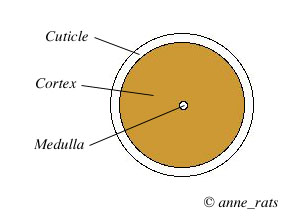
The hair shaft consists of three layers. The outermost layer is called the cuticle and consists of overlapping cells that look like shingles pointing upward and outward. They lock with the inner root sheath and firmly anchor the hair in the follicle. The cuticle also protects the soft inside of the hair shaft from damage.
The middle layer of the hair shaft is called the hair cortex. The cortex takes up the bulk of the hair. It consists of keratinized cells (cells that have become horny and impregnated with the protein keratin). The cortex also contains the granules of pigment that give the hair its color (for more on rat pigmentation, see Coat Color Mutations).
The innermost layer is the medulla. It forms the tiny core of the hair but is not always present, especially in fine hair.
A mutation called shorn may involve an error in the gene that codes for keratin, which leads to hairlessness (ongoing research, T. King, J. Mulrooney, R. Rollin, and K. Martin-Troy).
The Charles River hairless and fuzzy mutations in rats cause improper keratinization of the hair, combined with a thick layer of skin, which also produce hairlessness (Panteleyev and Christiano 2001).
Rats have several types of hair (Dry 1928):
More on hair:
|
|
|
Anatomy of a hair follicle |
Cells around the hair shaft itself are called the inner root sheath. The inner root sheath is embedded in the outer root sheath which is continuous with the epidermis.
The sebaceous gland produces oils which coat the hair.
All follicles are supplied with blood, a few nerve endings, and some tiny muscles responsible for piloerection in animals (and goosebumps in humans).
Hair follicle growth cycle
|
|
|
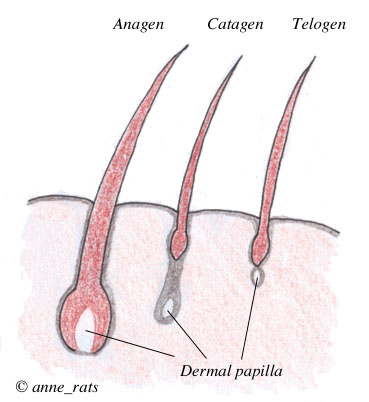
Hair follicle growth cycle |
The phase of active hair growth is called the anagen phase. Anagen phases may vary in duration, and the rate of growth during the anagen phase may differ. Guard hairs, for example, have a faster growth rate than down hairs, and they end up longer. Hairs produced in the anagen phase have thick shafts and are more deeply pigmented.
The rexoid mutations slow the rate of growth during the anagen phase, which results in shorter hair. The angora mutation does not increase the rate of hair growth, but lengthens the anagen phase itself, resulting in longer hair. The angora mutation has not yet been found in rats, however.
The anagen phase is very metabolically demanding, and is thus very sensitive to levels of thyroid hormone, a hormone that has a global influence on cellular metabolism. If thyroid hormone levels are too low, follicles can go into the resting phase prematurely, leading to loss of hair. The hairless (hr) mutation disrupts the action of thyroid hormone in humans and mice, leading to a lack of hair. The hairless (hr) mutation has not yet been found in rats, however.
After the anagen phase, the follicle starts to shut down. This is called the catagen phase. Cell division stops, the dermal palilla condenses, the epidermal sheath shrinks and is keratinized into a tiny "club root" attached to the base of the hair shaft, which is pushed upward toward the surface by epithelial cells.
After the follicle shuts down, the follicle enters the telogen phase, a resting phase during which the hair is free to fall out.
|
|
|
Cricket showing hair growth patterns |
In rats, hair follicles don't all follow the same cycle at the same time. Instead, the anagen growth phase passes like a wave from nose to rump, and from belly to back. Sometimes, during a rat's molt from one coat to the next, you can see funny coat patterns particularly on the rat's face or forehead, formed by the interface between the new and old coats.
Cessation of growth happens the same way, the catagen and telogen phases passing in a wave from nose to rump and belly to back. (In contrast, humans have a mosaic pattern of activity).
|
|
|
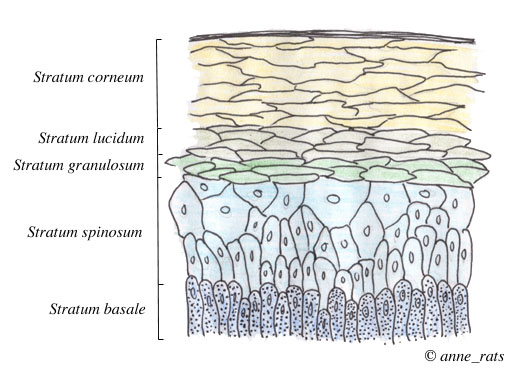
Cross section of the epidermis |
The Charles River hairless and fuzzy (fz) mutations in the rat causes a thick buildup of corneocytes. The hair shaft, which is improperly keratinized, cannot penetrate the thick stratum corneum. The hair follicle therefore fills with keratin, and may become keratin-filled cyst. The ultimate outcome is that the rat is hairless (Panteleyev and Christiano 2001).
Skin is a subcategory of epithelial tissue, one of the primary types of tissue in the body. There are two types of epithelium:
The differentiation of the epithelium into different tissues and the ongoing renewal of skin depend on a complex cascade of cellular events. If a mutation occurs in that cascade, the effects may be very diverse, from hair to skin to glands. An example of such a mutation is the nude mutation in rats (Rowlett nude (rnu), New Zealand nude (nznu)), which leads to skin anomalies, hairlessness, failure to develop a thymus, lactation problems, kidney failure and so forth.
A mutation in epithelial cell differentiation: the nude locus ( Rowlett nude rnu and New Zealand nude nznu)
|
|
|
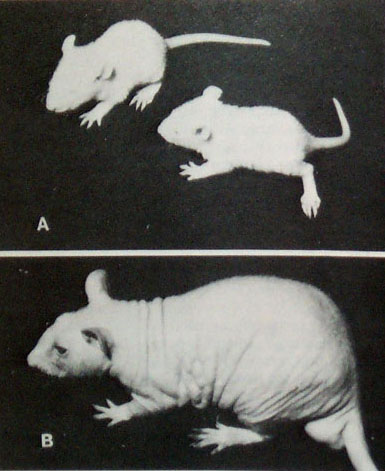
Homozygous nude (rnu/rnu) rat. Festing et al. 1978 |
Other effects: nude rats have no functioning thymus. Instead they have brown fat tissue and a small and highly abnormal thymus rudiment. The lack of the thymus leads to immunodeficiency (no cell mediated immune response e.g. T-cells). Nude rats grow at only 60-80% of the rate of normal rats (Festing et al. 1978).
Genetics: The nude locus codes for a protein called wing-helix protein, whn. Whn is involved in epithelial differentiation. It is required for normal development of the skin, hair and thymus. In the skin, Wing-helix is expressed in the epithelial cells. A defective wing-helix protein causes abnormal hair development.
Whn appears to mediate cell differentiation and transition. For example, cells at the base of the hair multiply rapidly. Then some of them stop multiplying and join the growing column of the hair shaft. Whn mediates the transition from the proliferative to the non-proliferative state of the cells in the hair base (Prowse et al. 1999).
Whn is a key regulatory factor which maintains the balance between keratinocyte growth and differentiation in the skin (Brissette et al. 1996, Baxter and Brissette 2002). Specifically, it oversees the initiation of terminal differentiation of keratinocytes into dead corneocytes (Lee et al. 1999). The rnu mutation leads to abnormal keratinocyte differentiation, and a super-sensitivity of keratinocytes to growth inhibition commands (Brissette et al. 1996, Baxter and Brissette 2002).
Whn is also implicated in the development of the thymus, a gland that is derived from epithelial tissue. Whn is required for the primordial thymus tissue to differentiate into a fully-formed normal thymus (Lee et al. 1999).
If the whn protein is disrupted, as it is in the Rowlett nude (rnu) mutation in rats, the primordial cells of the thymus fail to differentiate into the different mature cell types of the normal thymus. Nude rats therefore do not develop a thymus (Festing et al. 1978, Balciunaite et al. 2002).
T-lymphocytes mature in the thymus, so without a thymus one large branch of the immune system is non-functioning. Nude rats experience immune deficiency. Immune function that does not depend on the thymus (killer and natural killer cells) appears to be normal (Chassoux et al 1983).
A mutation that takes out three genes at once: two
for hair, one for kidney function: the shorn locus
(shn)
|
|
|
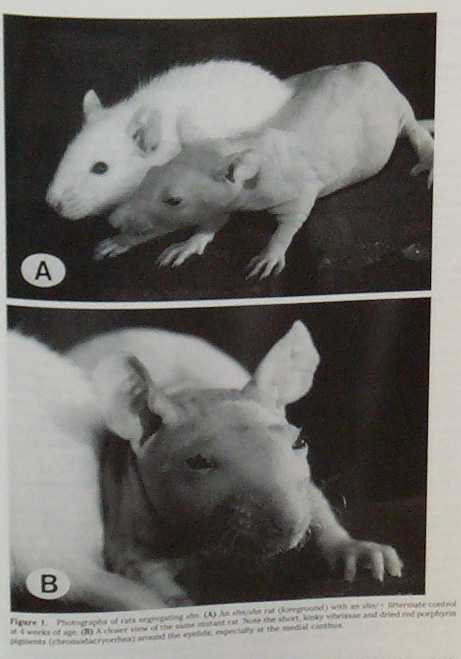
Homozygous shorn (shn/shn) at next to a heterozygous shn/+ littermate at 4 weeks. Moemeka et al. 1998. |
Other effects: Shorn rats develop abnormal hearts and kidneys which are yellow and bumpy instead of red and smooth. They tend to be smaller than normal rats and they die prematurely, at around 12-14 months of age (Moemeka et al. 1998).
Genetics: The shorn locus has been mapped to a narrow area of Chromosome 7 (Hall et al. 2000, Chrissluis 2002), which includes a number of genes: a Hoxc gene complex, keratin type II (krt2), and aquaporins 2 and 5. The Hoxc gene complex is responsible for normal hair development in mice (Godwin and Capecchi, 1998), and krt2 encodes the basic keratins, the major protein component of mammalian hair. Aquaporins are facilitate water transport across the cell membrane, and are found extensively in the kidneys (ongoing research, T. King, J. Mulrooney, R. Rollin, and K. Martin-Troy).
The shorn mutation may be a large mutation, such as an inversion, that impairs all three of these gene complexes at once.
Human analogues: Mutations in Aquaporins figure prominently in diabetes insipidus. People with diabetes insipidus cannot reabsorb water to produce concentrated urine, even if they are very dehydrated.
|
|
|
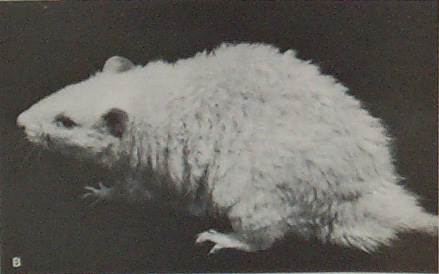
Top: two 15-day old fuzzy rats. Bottom: young adult male fuzzy rat (Palm and Ferguson 1976) |
Other effects:
- Kidney failure: Fuzzy rats suffered from chronic, progressive nephrosis, or nephropathy, which shortened their lifespan to 16.6 months in males and 20.4 months in females.
Details: Chronic, progressive nephrosis means the thickening of the basement membrane of the kidney with protein cases, and it leads to kidney failure. Chronic nephrosis progresses differently in male and female fuzzy rats. Generally, males are more affected, at an earlier age, than females are, though by old age all fuzzy rats had the condition. Specifically, chronic nephrosis started around 8 weeks of age in both sexes, but neither sex was obviously affected for several months. However, at 21 weeks of age almost all the males had mild nephropathy, which became diffuse nephropathy in of all the males by 32 weeks of age. Females remained healthier for longer, as they did not show diffuse nephropathy until 52 weeks of age, and even then it was in only half the females. By old age, however, all rats had progressive nephrosis.
Longevity: In one longitudinal study of the health of fuzzy rats all males and 80% of the females were debilitated by kidney failure and were eventually euthanized. Males averaged 16.6 months (range: 12-20 months), and females averaged 20.4 months (range: 17-26 months) at time of death. This age at death is younger than normal rats, indicating that fuzzy rats have a decreased lifespan (Marit et al. 1995).
- Tumors: The tumor incidence in fuzzy rats was similar to that in normal rats (70% females and 25% males developed tumors, almost all of which were benign), but fuzzy rats developed their tumors at an earlier age than normal rats (Marit et al. 1995).
- Adrenal glands: cystic degeneration of the adrenal gland (replacement of cortical cells by cyst-like spaces containing blood or serum). Females are more affected than males. This condition was rare and mild in young fuzzy rats, but by old age 100% of females had moderate to severe lesions, and 35% of males mild and a few moderate lesions (Marit et al. 1995).
- Skin: The skin of fuzzy rats has fewer, smaller, poorly developed follicles. Frequently the lumen of the follicle is enlarged and completely filled with keratin. These accumulations of keratin form cysts in the skin. The skin of fuzzy rats has a slightly thicker stratum corneum (Palm and Ferguson 1976, Ferguson et al. 1979, Marit et al. 1995).
- Hair: Hair shafts of fuzzy rats are smaller and do not contain the medulla. The corniform layer is slightly thicker and more loosely arranged than normal. Sebaceous glands are more prominent (Palm and Ferguson 1976, Ferguson et al. 1979, Marit et al. 1995).
- Other: Mild myocardial degeneration (cardiomyopathy), retinal atrophy (Marit et al. 1995).
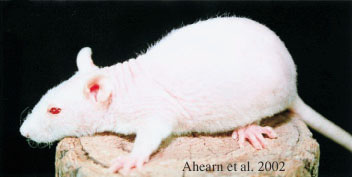
Genetics: The exact gene that is affected by the fuzzy mutation is not known. However, the fuzzy mutation has been mapped to a location on Chromosome 1 in the rat, and there are many good potential gene candidates at that spot. One of the most compelling genes that may be involved is the fibroblast growth factor receptor 2 gene (Fgfr2) which is expressed in hair follicles (Ahearn et al. 2002).
An allele of fuzzy: Charles River "hairless"
Appearance: Charles River "hairless" rats have abnormal keratinization of the hair shaft, and the formation of a thick, dense layer of dead skin cells (corneocytes) in the skin. This thick layer prevents the poorly keratinized hair shaft from penetrating to the skin surface. The hair follicles degenerate over just a few days, and by day 12 the skin has mostly degenerated hair follicles with abnormal or broken hairs.
|
|
|
Photo of a "hybrid" rat with one Charles River hairless and one fuzzy allele. (from: Ahearn et al. 2002) |
Genetics: The Charles river hairless mutation annuls cell proliferation in the hair matrix and affects keratinocyte differentiation in the hair follicle and in the skin (Panteleyev and Christiano 2001, Ahearn et al. 2002).
The Charles River "hairless" mutation is actually an allele of fuzzy in the rat on Chromosome 1, and frizzy in the mouse (Ahearn et al. 2002). There appear to be several mutations in the fuzzy gene in rats (Ferguson et al. 1979).
The so-called hairless mutation in the rat, which has been designated hr in the literature, is distinct from the mouse hairless mutation. The rat hairless mutation is actually the Charles River hairless, which is an allele of fuzzy (Ahearn et al. 2002, Panteleyev and Christiano 2001).
This fuzzy gene in the rat is an orthologue of the mouse frizzy gene. To make things even more complicated, mice also have a gene called fuzzy, but it isn't related to the rat's fuzzy.
Mouse hairless (the real hr) has been very well studied. Humans can have the hr mutation as well. Rats, however, do not have this hairless mutation. I've written up a summary of how mouse hr works, but remember that rats don't have this mutation.
This is all rather confusing, so here's a table showing the correspondence between the different mutations in rats and mice:
|
|
|
|
fuzzy (fz), Charles River hairless (hr) |
frizzy (fr) |
|
Rowlett Nude (rnu), New Zealand Nude (nznu) |
nude (Whn) |
|
shorn (shn) |
-- none -- |
|
-- none -- |
fuzzy (fz) |
|
-- none -- |
hairless (hr) |
Slowing the rate of hair growth: the rexoid mutations (rex and wavy)
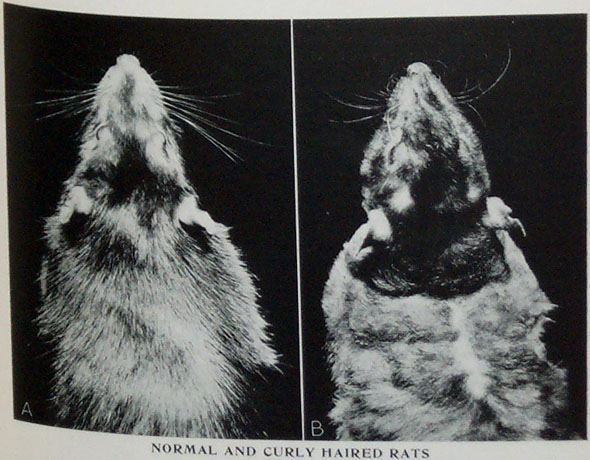
The rexoid mutations slow the rate of hair growth. The end result is that many or all of the hairs are shorter than usual. The hairs are also thin and imperfectly formed. The coat presents with a characteristically curly appearance.
The rexoid mutations affect all hair types, but especially the guard hairs. The guard hairs still grow a little faster than wool hairs, but the difference is slight and the end result is that the guard hairs aren't much longer than the wool hairs. Guard hairs still have their coarse tip, but it isn't as coarse or as long as in normal guard hairs (Fraser 1953, Robinson 1981).
Currently, seven genes are known to produce a rexoid phenotype: wavy, rex, ragged, curly-1, curly-2, kinky, and shaggy. However, the last four mutations (curly-1, curly-2, kinky, and shaggy) appear to be extinct in laboratory rats (Greaves 1981). Ragged is rare, as it was only recently discovered and developed in Japan.
The frequency of these different mutations in pet rats is unknown. Pet rats may also have additional rexoid mutations that have not yet been described or studied.
Wavy
|
|
|
Homozygous wavy (wv/wv) rat. Greaves 1981 |
Pelage of wavy rats (wvwv) is distinctly wavy or curly, most of the hairs have 1-6 waves or kinks along their length. The whiskers are mostly wavy or bent, are shorter and thinner than normal whiskers. Coat is thinner and less curly than rex rats, though wavy rats and rex rats can be hard to tell apart. Wavy rats have normal fertility and a normal lifespan (Greaves 1981, Robinson 1988).
Rex
|
|
|
Heterozygous rex (Re/+) rat. Greaves 1981 |
The pelage of rex heterozygotes (Rere, known as rex) appears wooly and unkempt. The coat starts out soft in very young rats, and a rough texture develops through molts in adolescence and adulthood. Rex rats have a good covering of hair but the skin may be seen with each movement (Robinson 1981).
Homozygotes (ReRe, known as patchwork, semi-hairless, or double-rex) develop just like rex rats until about four weeks of age, when the coat is rapidly lost. Hair loss starts on the head and shoulders and spreads to the rest of the body (not surprisingly -- it follows the wave of hair follicle cycling from nose to tail). Hair loss is usually complete by day 36. A new coat erupts at each molt, but subsequent coat is thin, with some bareness. Their hairs are easier to pluck out than in normal rats and rex rats. Females are more affected than males and have larger bald areas (Robinson 1981).
Rex vs. Wavy
Rex and wavy are not allelic -- they are not alleles of the same gene, but are mutations of separate genes. Rex coats tend to be thicker and more curly than those of wavy rats, though the two types can be hard to distinguish reliably. Rex and Wavy rats can be bred together. Rats that are Rex-Wavy (Re+/wvwv) have fur that is exceptionally sparse, amounting to little more than a thin, downy covering over much of the body (Greaves 1981).
Ragged was first reported in Japan in 1990. Ragged rats have a ragged coat by age 10 days, then lose most of the hair on their backs by 5 weeks of age. After 5 weeks, they grow a new coat of ragged hair. Ragged rats also develop thickened eyelids, and have thicker and more numerous sebaceous glands in the skin of the back. Ragged is a single, recessive trait (rg) (Noguchi 1990).
In 1981 four additional rexoid variants were known in Norway rats (Robinson 1981, Greaves 1981). These four mutations are currently thought to be extinct:
- Guard hairs and whiskers are wavy and somewhat curled. The curl becomes somewhat indistinct during early adulthood (between 30 and 200 days). Coat is softer than usual. See picture under "Shaggy" for photograph of Curly-1 rat.
General appearance very similar to Cu-1, but curl is stronger. Young develop short, crooked or curly whiskers that curve toward the back. Adults have fewer, smaller guard hairs which are distinctly curly. Underfur shows through. Appearance is curly and rough. Homozygotes, Cu-2/Cu-2 look like heterozygotes Cu-2/cu-2, indicating true dominance.
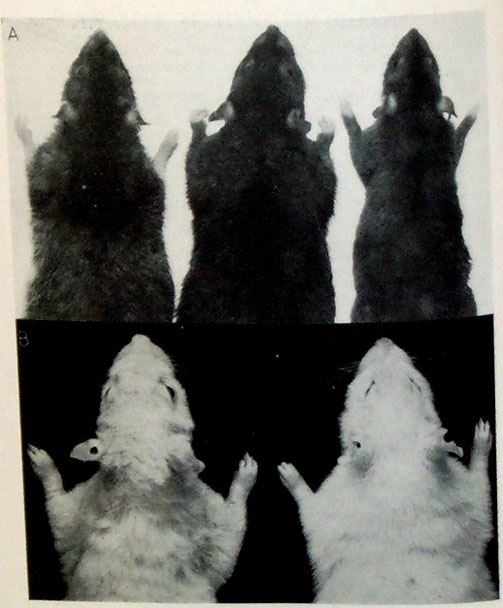
Normal (left) and Curly-2 (right) rats (Gregory and Blunn 1936)
|
|
|
Normal (left) and kinky (k/k) (right) rats. Feldman 1935. |
- Young develop curly or kinky whiskers and coat. The kinky-hair curl is more pronounced and more consistent over the lifespan than the curl of Cu-1-haired rats. Kinky juvenile coat is shorter than normal, appears very dense, and feels wooly. Adult pelage is shorter and rougher than usual. Hairs are flattened. Over six months of age, new hair growth does not keep pace with an excessive breakage of hair, leading to bare spots (particularly on the mobile areas of shoulders and hips). Coat does not keep them as warm as normal rats. Kinky-haired rats are smaller, have reduced fertility, and a shorter lifespan than normal. Kinky is not an allele of Cu-1.
Effect on hair is intermediate between Cu-1 and Cu-2. Closely linked to Cu-1 on chromosome II. Rats with both Curly-1 and Shaggy are more curly than a rat with just Curly-1 or kinky, indicating interaction between the two genes.
Top row: Curly-2, Curly-1, and Shaggy (all heterozygotes). Bottom row: An F1 Shaggy/Curly-1 (left), a homozygous kinky rat (right)
How are rex and wavy related to curly-1, curly-2, kinky and shaggy?
Unfortunately, the four variants curly-1, curly-2, kinky and shaggy appear to be extinct. Nobody knows for sure how rex and wavy are related to these four mutations. Rex and wavy could be the same mutation as one of these, they could be alleles of one of these four (a different mutation in the same gene), or they could be new mutations. Here are some ideas about the relationship between rex and wavy and these four earlier mutations as reported in the literature (Robinson 1981, Greaves 1981):
Other coat types in domestic rats
There are other rat hair types out there, such as "velour," "velveteen," "velvet," and "teddy rex." These mutations may represent re-occurrences of Cu-1, Cu-2, kinky or shaggy, they may be alleles of these genes, or these mutations may be new. Their genetic underpinnings are speculative at this point.